
Download
Journal of Biological Methods | 2023 | Vol. 10 | e99010006
DOI: 10.14440/jbm.2023.410
DOI: 10.14440/jbm.2023.410
ARTICLE
Optimizing factors for large-scale production of Arbuscular Mycorrhizal Fungi consortia using root organ cultures
Maunata Ghorui1, Shouvik Chowdhury1, Keshab Das2, Kiran Sunar2*, Balu Prakash1*
1Department of Biotechnology, School of Life Sciences, Vels Institute of Science, Technology and Advanced Studies (VISTAS), Pallavaram, Chennai - 600117, India
2Department of Botany, Balurghat Mahila Mahavidyalaya, Balurghat, Dakshin Dinajpur, West Bengal – 733101, India
2Department of Botany, Balurghat Mahila Mahavidyalaya, Balurghat, Dakshin Dinajpur, West Bengal – 733101, India
*Corresponding authors: Dr. Balu Prakash, Department of Biotechnology, School of Life Sciences, Vels Institute of Science, Technology and Advanced Studies (VISTAS), Pallavaram, Chennai - 600 117, India, prakazbt@gmail.com; Dr Kiran Sunar, Department of Botany, Balurghat Mahila Mahavidyalaya, Balurghat, Dakshin Dinajpur, West Bengal – 733101, India, kiran.sunar@gmail.com
Competing interests: The authors have declared that no competing interests exist.
Abbreviation used: AMF: Arbuscular Mycorrhizal Fungi, ROC: Root Organ Culture, DW g L-1: Dry weight gram per litre of media, ppg L-1: Propagules per liter of media, MTT: (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, RH: Relative Humidity
Received August 24, 2023; Revision received October 11, 2023; Accepted October 17, 2023; Published November 8, 2023
Abstract
Large-scale production of Arbuscular Mycorrhizal Fungi (AMF) consortia is a crucial stride in harnessing their potential for sustainable agriculture and plant growth enhancement. However, establishing optimal production conditions is challenging due to their obligate nature, variability, lack of standardized protocols, and limited understanding of their specific requirements. Previous attempts to standardize Root Organ Cultures (ROC) for AMF overlooked challenges related to viable inoculum production for field applications. This current investigation reported, for the first time, the optimization of various factors during large-scale production of AMF using ROC. By optimizing factors like gelling agents, media preparation, medium-to-inoculum ratios, incubation conditions, age, harvesting method and drying temperatures, we achieved significant yields of viable propagules. The standardized protocol outlined in this study will greatly influence commercial-scale AMF production. These standardized protocols are poised to contribute to larger-scale AMF production worldwide, with the potential to support sustainable agriculture and ecosystem management.
Keywords: Arbuscular Mycorrhizal Fungi (AMF), Large-scale production, Root Organ Culture (ROC), Yield, Spore, Culture, Media, Propagule
1.BACKGROUND
Arbuscular mycorrhizal fungi (AMF), which are involved in a symbiotic relationship between plants and fungi, play a crucial role in enhancing plant nutrient uptake, improving soil structure, and promoting overall growth and health of plants [1]. The availability and effectiveness of AMF inoculum are of utmost importance to the successful establishment and functioning of this symbiosis. In vitro techniques have emerged as a viable option for large-scale multiplication of AMF inoculum thanks to their ability to provide large quantities of sterile inoculum within a controlled environment and a relatively short time [2,3]. Indeed, as reported, in vitro monoxenic cultivation techniques are now a practical reality, providing cost-effective alternatives to chemical fertilizers. To enhance their economic attractiveness, adjustments are required, such as identifying limitations, employing affordable, eco-friendly inputs, and optimizing production scale [4]. Among various in vitro mass multiplication techniques, multiplication using root organs on solidified medium, known as Root Organ Culture (ROC), stands out [3]. The controlled environment of ROC ensures the absence of soil-borne pathogens and contaminants, resulting in the production of sterile AMF inoculum [5]. Large-scale cultivation of AMF using root organs on solidified medium, like other mass-multiplication techniques, is subject to several crucial factors that determine the infectivity potential of the inoculum when introduced into the soil. These factors include the choice of gelling agents, starter culture biomass, ageing duration, incubation conditions, and harvesting methodology. Understanding the influence of these in vitro manufacturing factors is crucial to the optimization of the infectivity potential of AMF inoculum, which ultimately leads to improved plant performance, increased crop yield, and enhanced soil health. Though there are various reports on and methodologies for the in vitro cultivation of AMF using ROCs, the evidence supporting consistent and high-quality AMF production remains limited. While root organ cultures have been utilized for many years for the cultivation of AMF, there hasn't been a universally-accepted method for evaluating their effectiveness in propagating these fungal symbionts [6]. In view of this critical need, this study aimed to define a set of standardized fundamental parameters that would exert a positive influence on both quality and quantity of AMF production through ROC.
2.MATERIALS AND METHODS
Our research complied with the foundational guidelines set forth by various researchers over time, including Adholeya [7], Bécard & Fortin [8], Sandel [9], Doner & Bécard [10], Sylvia & Jarstfer [11], An et al. [12], Phillips & Hayman [13], and others. However, these existing protocols lacked details specifying essential aspects such as choice of gelling agents, pH levels, sub-culturing ratios, incubation conditions, harvesting techniques and post-harvesting inoculum processing procedures. Considering these gaps in knowledge, our study extensively investigated the impact of these factors on various dependent variables. These included physicochemical attributes of the growth medium, root biomass production, propagule yield, spore generation, spore viability, root colonization, contamination, and propagule recovery.
2.1. Culture Media
2.1.1. M-media composition
The composition of the M-media used in our study was based on the formulation introduced by Bécard & Fortin in 1988 [8], with some modifications in the gelling agent and its concentration. While the original authors utilized 0.3% Bacto Agar, we adjusted and compared 0.23% Gellan gum and 0.35% Phytagel, used as the gelling agent in our experiments. Medium composition is detailed in Table 1.
2.1.2. Physicochemical characteristics of Gelling Agents
In our study, Gellan gum (0.23% w/v) and Phytagel (0.35%) were used and compared. The media characteristics to be evaluated were adapted from American Pharmaceutical Review [9]. Physical tests included visual test for colour, clarity, gel strength, pH, and damage checks, such as breaking of the gel while handling the culture media.
2.1.3. Sterilization
In our research, we used both the conventional autoclave as well as an automated media maker for media production. Both types of equipment were tested for their productivity and their influence on propagules recovery.
Table 1. Composition of M-Medium based on the formulation of Bécard & Fortin in 1988 [6]
| Chemicals | mg L-1 |
|---|---|
| MgSO4.7H2O | 731 |
| KNO3 | 80 |
| KCl | 65 |
| KH2PO4 | 4.8 |
| Ca(NO3)2.4H2O | 288 |
| MnCl2.4H2O | 6 |
| ZnSO4.7H2O | 2.65 |
| H3BO3 | 1.5 |
| CuSO4.5H2O | 0.13 |
| Na2Mo4.2H2O | 0.0024 |
| NaFeEDTA | 8 |
| KI | 0.75 |
| Glycine | 3 |
| Thiamine HCl | 0.1 |
| Pyroxine | 0.1 |
| Nicotinic Acid | 0.5 |
| Myoinositol | 50 |
| Sucrose | 10000 |
The medium pH was adjusted to 5.5 before sterilization.
2.2. Inoculation conditions
Sub-culturing or inoculation involved transferring a culture inoculum from a pre-maintained starter culture unit to a new culture medium. The cultures were incubated under specific environmental conditions to promote growth till the desired level of growth has been attained, which was followed by the harvesting of the cultures.
2.2.1. AMF species
The root organ culture for AMF consortia was obtained from Symbiotic Sciences Pvt Ltd – ROC of Rhizophagus species (SSDP005).
2.2.2. Host
Genetically-modified roots of Daucus carota
2.2.3. Temperature
~25oC
2.2.4. Light conditions
Dark
2.2.5. Inoculum Starter
In this study, two different quantities of basic inoculum (Starter Culture) were used: 0.13 DW g L-1 and 0.20 DW g L-1. The purpose was to compare the weight of the two starter cultures to ascertain which of them yielded the most significant production outcomes.
2.2.6. Humidity
The study focused on investigating the influence of ~50% and ~60% relative humidity on contamination levels.
2.2.7. Duration
Cultures were subjected to ageing ranging from 66 to 90 days to identify the optimal age for harvesting that resulted in maximum yields.
2.3. Harvesting
2.3.1. Culture harvesting method
The cultures were harvested using a chelation solubilization method described by Doner & Bécard, 1991 [10], which involved exposing the gellan to 10 mM sodium citrate buffer (pH 6.0). This approach allowed for the recovery of viable plant tissues and AMF propagules from the culture while deionizing the gelling agent. The recovered biomass and propagules were subjected to washing to eliminate any remaining chemical residues.
2.3.2. Harvesting equipment
In this study, two types of equipment were utilized, i.e., an orbital shaker and a liquid blender. The liquid blending method, which involves high sheer size reduction in a liquid as described by Sylvia & Jarstfer [11], was employed. The study focused on evaluating the productivity of both equipment types and their influence on propagule recovery.
2.3.3. Biomass drying
The biomass was subjected to drying at ~25°C (air-drying) for 60 hours and ~37°C for 36 hours, and the study examined the influence of these drying conditions on the yields.
2.3.4. Root processing
The roots were subjected to milling and sifting by using automated grinder to get the propagules sized >63 µ.
2.4. Post-harvesting evaluations
2.4.1. Root biomass yield (g L-1)
Harvested roots were washed, dried. The root biomass yield was calculated by dividing dry biomass (g) by volume of media (L-1) and expressed as weight per volume of media (g L-1).
g L-1 = Dry biomass/Volume of media
2.4.2. Propagule yield (ppg L-1)
Both the spores and root fragments were considered as propagules. Propagule yield was determined by counting the propagules in 500 µL aliquots taken from 0.01 g of inoculum hydrated in 20 mL of distilled water and the determination was done in triplicate for each sample. The formula for calculating propagules per gram (ppg g-1) and propagule yield (ppg L-1) were:
ppg g-1 = [(Spores + Root Fragments) * (1000 µL/500 µL) * 20mL] / 0.01 g
ppg L-1 = Propagule per gram (ppg g-1) * Root biomass yield (g L-1)
2.4.3. Spore yield (sp. L-1)
To calculate the spore yield, the same methodology used for the calculation of propagule yield was employed, but only spores were counted. The formula for calculating spores per gram (sp. g-1) and spore yield (sp. L-1) of the inoculum was as follows:
sp. g-1 = (Spores) * (1000 µL/500 µL) * 20mL] / 0.01 g
sp. L-1 = Spore per gram (sp. g-1) * Root biomass yield (g L-1)
2.4.4. Spore viability (%)
The method described by An et al. [12] was employed for the calculation of spore viability, with some modifications. In the original study, incubation duration was 40 hours with no temperature specified. In our study, the time was extended to 48 hours and the temperature was set at 37oC. 100 randomly collected spores were placed in 30-mm Petri dishes with three replicates set for each sample. Spore suspensions were diluted with the stock of 0.5 mg MTT mL-1 (Himedia, CAS No. 298-93-1) at a ratio of 1:1 and incubated at 37oC for 48 hours. Viable spores appeared pink or red, while non-viable ones looked green, dark, or unstained under yellow light. The spore viability percentage was calculated using the formula:
Spore viability (%) = (Number of spores stained red or pink /Total number of spores) * 100
2.4.5. Clearing of roots, staining, and scoring the Mycorrhizae
The technique was based on the method proposed by Phillips & Hayman [13] with modifications. They heated root fragments at 90°C for 1 hour in 10% potassium hydroxide, acidified with dilute hydrochloric acid, and stained by simmering for 5 minutes in 0.05% Trypan Blue. Our method involved autoclaving ~5 mg inoculum with 10% potassium hydroxide for 15 minutes, washing, treating with 10% hydrochloric acid for 30 mins, and autoclaving with 0.05% Trypan Blue for 15 minutes. Mycorrhizal colonization was assessed by scoring its presence or absence using a specific formula:
Root colonization (%) = (Number of colonized root fragments /Total root fragments observed) * 100
2.4.6. Propagule recovery
It refers to the viable portion of the sample that remains after removing the contamination. It is calculated as follows:
Propagule recovery = (1 – Contamination %) * Propagule yield (ppg L-1)
2.4.7. Contamination (%)
Contamination was detected through visual examination. Bacterial contamination presents as cloudiness or turbidity, and sometimes a thin film on the culture's surface. Mold contamination manifests as the presence of multicellular filaments or mycelium. Yeast contamination results in turbidity in more advanced stages. Any such changes indicated contamination [14]. It was calculated by:
Contamination (%) = (Contaminated media or cultures / Total media or cultures) * 100
2.4.8. Equipment productivity
Equipment productivity refers to the number of media or cultures processed by the equipment in each unit of time.
Equipment & methodology Productivity = Number of media or cultures processed / Total duration of equipment usage
2.5. Statistical analysis
The statistical analysis was conducted using R version 4.3.0 software [15]. A Welsch t-test was used to compare groups with a 95% confidence level (alpha = 0.05), to determine significant differences between the groups. Regression analysis was also performed to examine the impact of predictor variables on the response variable, assessing relationships and factors influencing the response variable. Additionally, the Pearson correlation coefficient was calculated to assess the strength and nature of linear relationships between pairs of variables.
3. RESULTS
3.1. Culture Media
3.1.1. Physicochemical analysis of the gelling agents
Table 2 presents a comprehensive comparison of the physicochemical characteristics of two gelling agents, Phytagel and Gellan gum. Gellan gum required a lower concentration (0.23% w/v) than Phytagel (0.35% w/v) to achieve similar gel strength. Both gelling agents yielded transparent and clear media upon gel formation, with impurities occasionally observed in Phytagel. Gellan gum maintained a firm gel structure, while Phytagel exhibited relatively weaker gel strength. Gellan gum demonstrated a slightly lower pH (5.6±0.1) compared to Phytagel (5.8±0.2) before pH adjustment. Phytagel occasionally showed susceptibility to cracking during handling, while Gellan gum remained structurally intact. Phytagel occupied more space with a lower bulk yield, different from Gellan gum with a higher bulk yield. Syneresis was more pronounced with Phytagel, causing gel shrinkage and liquid separation, whereas Gellan gum exhibited better retention of its gel structure with minor syneresis. Furthermore, Phytagel was associated with quicker oxidative browning of roots, while Gellan gum displayed slower browning. These disparities could potentially guide the selection of a gelling agent based on considerations of space efficiency, syneresis control, and prevention of oxidative browning within the culture system.
Table 2. Physicochemical characteristics of gelling agent
| Parameters | Phytagel | Gellan gum |
|---|---|---|
| Gelling concentration (w/v) | 0.35% | 0.23% |
| Color of media after gelling | Transparent | Transparent |
| Transmittance (based on COA) | 100% | 100% |
| Clarity | Clear, occasional impurities | Clear |
| Gel strength | Poor | Firm |
| Media pH before adjustment | ~5.8±0.2 | ~5.6±0.1 |
| pH adjustment required to reach 5.5 standard | Required adjustment | No adjustment required |
| pH 24-hrs post autoclave (Mean±SD) | 5.5±0.2 | 5.4±0.1 |
| Damage | Media crack while handling | None |
| Other characteristics | Low bulk yield Comparatively high syneretic Quicker oxidative browning of roots | High bulk yield Slight syneretic Slower oxidative browning of roots |
3.1.2. Identifying the impact of gelling agents on dependent variables
The impact of gelling agents was assessed using Welch Two Sample t-tests at a 95% confidence level, revealing different outcomes for Phytagel and Gellan gum. Contamination levels were significantly different (t = 5.521, p = 0.0001139) between Phytagel (mean = 29.5%) and Gellan gum (mean = 8.5%). Phytagel showed significantly lower propagule yield (t = -5.9791, p = 4.494e-05) and lower propagule recovery (t = -6.4097, p = 3.289e-05) than Gellan gum. However, no significant differences were found in Root biomass yield (Phytagel mean = 1.30, Gellan gum mean = 1.51) and Spore Yield (Phytagel mean = 177,036, Gellan gum mean = 290,224). These findings underscored the importance of choosing the appropriate gelling agent for specific desired outcomes.
3.1.3. Determining the optimal gelling agent for large-scale production
Gellan gum outperformed Phytagel by showing lower contamination (8.5% vs. 29.5%) and higher propagule yield and recovery. Propagule yield was found to be significantly lower with Phytagel (mean = 531,107) compared to Gellan gum (mean = 1,639,688) In terms of propagule recovery, Gellan gum displayed an average of 1,511,567 ppg L-1, whereas Phytagel exhibited a five-times lower average (377,413 g L-1). This analysis suggested that Gellan gum can be a potentially favorable gelling agent.
3.1.4. Media Preparator Productivity
Table 3 compares the the Autoclave and the Media Preparator for media preparation in terms of productivity. The Autoclave was characterized by a larger chamber volume, but it exhibited a lower productivity (20 L hr-1) when compared to the Media Preparator (34.7 L hr-1). The Media Preparator allowed for automated operations and quicker processing (3.45 hours). While the Autoclave was cost-effective, the Media Preparator proved to be more efficient and technologically advanced. The selection of equipment for media preparation was influenced by considerations of productivity requirements, ease of use, and budget limitations.
Table 3. Working capacities and specifications of Classical Autoclave and Media Preparator
| Factors | Autoclave | Media Preparator |
|---|---|---|
| Automated process | Heating, Sterilization | Heating, Sterilization, Cooling, Dispensing |
| Chamber Volume capacity | 810 | 125 |
| Maximum utilizable capacity | 120 | 120 |
| Time required from loading to dispensing at 45°C | 6hrs | 3.45hrs |
| Equipment & methodology productivity (L hr-1) | 20 | 34.7 |
| Ease of operations | Manual | Automated |
| Media dispensing per hour | 20 | 30 |
| Cost | Economic | Expensive |
3.2. Inoculation
3.2.1. Impact of medium-to-starter culture volume ratio on yields
Two different volumes of starter culture (0.13 DW g L-1 and 0.20 DW g L-1) were investigated for their influence on root biomass yield, spore yield, and colonization. The regression analysis revealed that the impact of starter culture biomass on these variables were not statistically significant (root biomass yield: p = 0.18694, spore yield: p = 0.592, colonization: p = 0.05641). The R-squared values indicated a low percentage of variability for root biomass (5.93%), spore yield (1.01%), and colonization (11.99%). So, starter culture of 0.13 DW g L-1 is preferred.
3.2.2. Impact of the relative humidity (RH) on culture contamination
The Welch Two Sample t-test was conducted to compare contamination levels between RH 60% and RH 50% and the result showed a significant difference in the level between the two groups, with a t-value of 10.153, df of 68.903, and a very low p-value of 2.522e-15. Contamination at 60% RH was 32.46%, while at 50% RH, it was 9.75%, indicating a higher contamination level at 60% RH.
3.2.3. Determining the optimal culture age to achieve the highest yield
Regression analyses demonstrated significant positive relationships between culture age and the root biomass yield (coefficient = 0.037446, p = 0.00885), propagule yield (coefficient = 55688, p = 0.00139), and spore yield (coefficient = 11419, p = 0.000864), being 70.77%, 83.92%, and 86.22% of their respective variations. However, Age did not bear a significant relationship with spore viability (coefficient = -0.14535, p = 0.104) and colonization (coefficient = 0.02717, p = 0.642), being 37.95% and 3.84% of their variations, respectively. It was observed that 85-88 days were optimal for achieving maximum the root biomass yield, propagule yield, and spore yield in in vitro cultivated AMF. The linear regression models for various dependent variables with respect to age are:
Root biomass = -2.090059 + 0.048290 * Age
Propagule density = -5624860 + 89222 * Age
Spore density = -494217 + 9466 * Age
Spore viability = 88.548624 + 0.001025 * Age
Colonization = 41.15109 + 0.04888 * Age
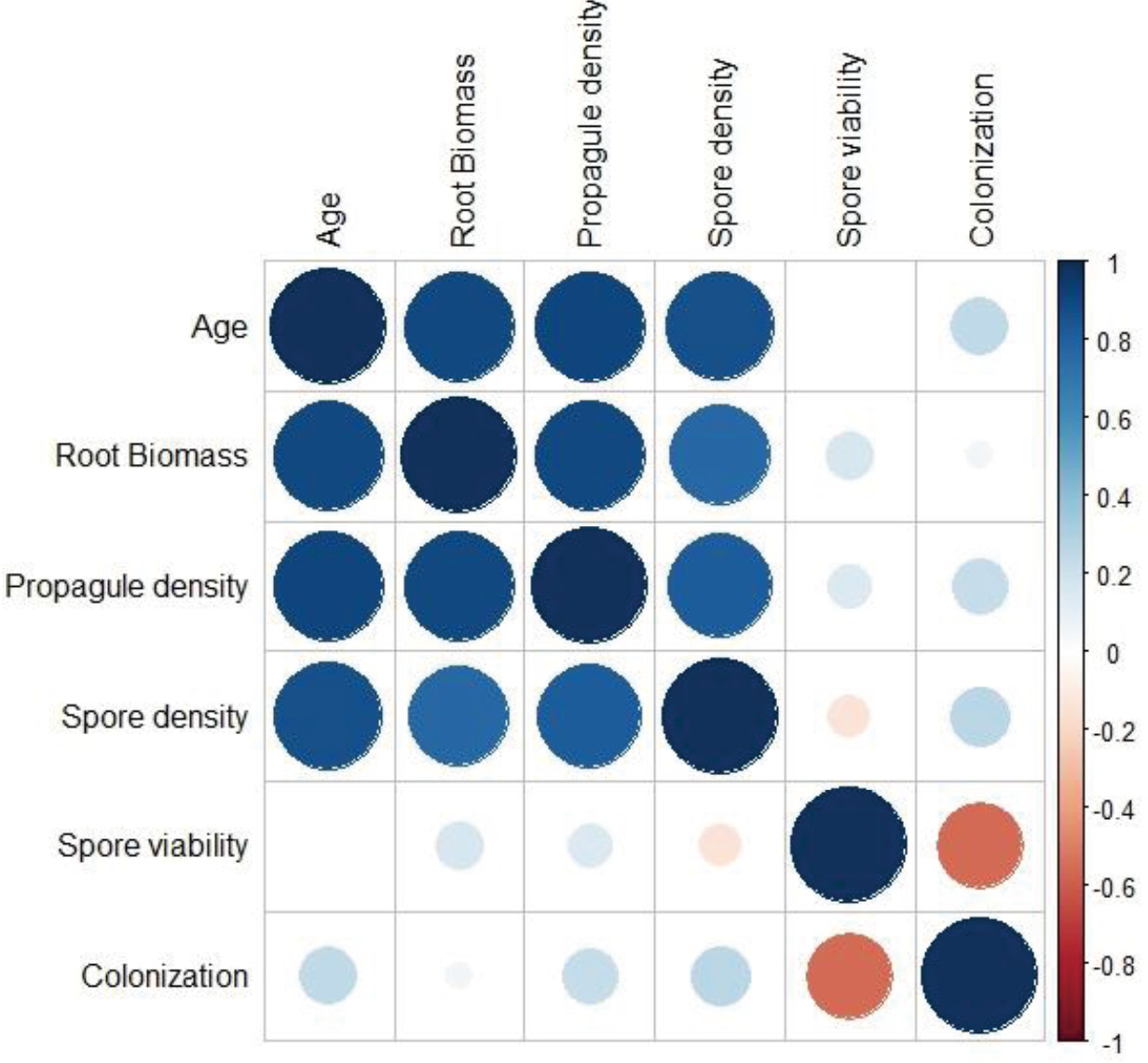
Figure 1 illustrates the strength and direction of the linear relationship between variables. There was a strong positive correlation between age and root biomass yield (0.84), propagule yield (0.92), and spore yield (0.93), and a moderate positive correlation between root biomass yield and propagule yield (0.69) and spore yield (0.71). Age exhibited a negative correlation with spore viability (-0.62), indicating an inverse relationship, while colonization did not show a significant correlation with other variables.
Figure 1 Correlation matrix showing the strength and direction of relationship between variables. Age has a strong positive correlation with Root Biomass (0.893), Propagule density (0.901), and a moderately positive correlation with Spore density (0.869) suggesting that these parameters tend to increase as age increases. Root Biomass has a positive correlation with Age (0.893), Propagule density (0.899), and a moderate positive correlation with Spore density (0.764) implying that Root Biomass is likely to increase alongside these variables. Propagule density has a positive correlation with Age (0.901), Root Biomass (0.899), and Spore density (0.812) implying that Propagule density tends to rise as these variables increase. Spore density has a moderate positive correlation with Propagule density (0.812) and a weak negative correlation with Spore viability (-0.130) suggesting that higher Spore density may coincide with higher Propagule density and lower Spore viability. Spore viability has a weak positive correlation with Root Biomass (0.168) and Propagule density (0.146), but these correlations are not strong. It has a negative correlation with Colonization (-0.552), suggesting that higher Spore viability might be linked to lower Colonization. Colonization has a moderate positive correlation with Spore density (0.270) and a strong negative correlation with Spore viability (-0.552). This implies that higher Colonization is associated with higher Spore density and lower Spore viability.
3.3. Harvesting
3.3.1. Harvesting equipment productivity
Table 4 makes a comparison between shaker and blender in terms of media deionization. While the shaker could handle larger volumes, it was more time-consuming. In contrast, the blender demonstrated a shorter deionization duration, greater productivity, and lower power consumption. The selection of equipment is subject to such factors as efficiency (time), productivity, and energy consumption.
Comparisons were made in terms of the effect of blender and shaker harvesting equipment on various yields and spore viability by using Welch Two-Sample t-tests. Significant differences were observed in root biomass yield and propagule yield. The blender group had higher mean root biomass yield (2.275 g L-1) and propagule yield (577,745 ppg L-1) compared to the shaker group (2.032 g L-1 and 506,489 ppg L-1, respectively). No significant differences were found in spore yield and spore viability. These results showed that blender had a higher productivity than shaker.
3.3.2. Biomass drying
Welch Two Sample t-test comparing spore yield when dried at ~25°C (air-drying) for 60 hours and ~37°C for 36 hours and the results showed a significant difference (p = 0.003061), with ~25°C group (133,606) having a higher mean yield than the ~37°C group (97,448). However, no significant difference (p = 0.6288) in spore viability was observed between the ~25°C (92.6%) and ~37°C (92.09%) groups. While drying at 25°C led to a higher spore yield, the trade-off was longer drying time. Therefore, one must consider the balance between maximizing spore production and optimizing the overall efficiency of the process.
3.4. Methodologies and Yields: Published Results vs. Our Findings
Table 5 presents a comprehensive comparison between methodologies employed in previous studies and our current investigation, in terms of various aspects of techniques, i.e., gelling agents, substrates, media compositions, inoculum sources, mycorrhizal fungi genera, host plants, incubation conditions, durations, and key outcomes withregard to spore count, propagule yield, biomass production, and colonization levels in the final product.
Earlier studies encompassed diverse approaches, including plant root organ cultures (ROCs), which utilizes porous substrates and nutrient solutions, classical ROCs with gellan gum or agar-vermiculite mixes, airlift bioreactors, Petri dish cultures, and ROCs in solid media with different gelling agents. The mycorrhizal fungi genera varied, including Rhizophagus, Gigaspora, Scutellospora, Sclerocystis, Acaulospora, and Funneliformis. Diverse host plants were used, including Daucus carota, Trifolium, chicory, clover, and bindweed.
Incubation conditions ranged from 24°C to 27°C, and the study duration varied from several weeks to months. Notably, spore and propagule yields displayed considerable variations, spanning from a few thousand to over two million spores per litre. Biomass production also showed significant variability, while colonization percentages ranged from 20% to over 75%.
In contrast, our present investigation utilized ROCs with Gellan gum as the gelling agent and M-medium as the substrate. The study achieved remarkable spore and propagule yields, biomass production, and colonization levels, contributing to the advancement of in vitro AMF propagation techniques. Figure 2 shows the outcome of current investigation, including enhanced sporulation during culture, viable spore counts and the finished final product.
Table 4. Working capacities and specifications of deionization methodologies
| Factors | Shaker | Blender |
|---|---|---|
| Cultures per round | 5L per carboy | 1L per blend |
| Preparation time-cleaning, filling (hours) | 1 | 0 |
| Deionization time (hours) | 4 | 0.5 |
| Total time (hours) | 5 | 0.5 |
| Sieving and washing roots (hours)-Simultaneous | 0.5 | 0.5 |
| Maximum media that can be harvested in 8 hrs | 64L | 640L |
| Equipment & methodology productivity (L hr-1) | 8 | 80 |
| Electrical consumption | 2 shakers for 4 hrs | 1 blender for 30 min |
| Unit consumed | 1.04 | 0.45 |
| Units consumed in 8 hrs (Watts) | 16.64 | 7.2 |
4. DISCUSSIONS
4.1. Culture Media
4.1.1. Gelling agent
Prior research revealed that solid media excelled in mycorrhizal colonization and arbuscule formation compared to liquid media. Immobilizing spores on solid media enhanced spore and hyphae production, while insufficient aeration in liquid media impeded the development of AMF [16]. The selection of gelling agents is crucial for in vitro cultivation of AMF, as they affect the medium's properties [17]. Different gelling agents, in terms of type, manufacturer, and concentration, have been shown to modify plant growth in vitro [18]. This study compared Gellan gum and Phytagel, both being bacterial polysaccharides produced from Sphingomonas elodea, and found that, as gelling agents, they are preferred to other alternatives such as agar or carrageenan. The use of Gellan gum and Phytagel, instead of Bacto-Agar, as described by original author Bécard & Fortin (1988) [8], was primarily driven by cost considerations. Gellan gum and Phytagel are significantly more cost-effective, being one-tenth of the price of Bacto-Agar [19]. When it comes to large-scale production, cost-effectiveness becomes a crucial factor. The objective is to achieve the best possible results with minimal input costs, including lower expenses for raw materials and reduced risk of contamination. The choice to lower the concentration of gellan gum to 0.23% was also motivated by the goal of cost reduction without compromising the final results. However, in the case of Phytagel, reducing the concentration to 0.3% proved inadequate in terms of the achievement of the desired viscosity, leading to increased syneresis, thus leading to contamination and reduced yields. As a result, 0.23% Gellan gum was compared with 0.35% Phytagel to find the optimal balance. Additionally, Gellan gum provided higher transparency for clear observation of root and tissue development and did not require any additional boiling before sterilization. We also found that it was mold-resistant, thereby facilitating early detection of contamination. It also exhibited higher heat resistance, withstanding temperatures up to 120°C, and solidifying within a range of 10°C-50°C as reported in previous studies [20],and showed minimal syneresis compared to agar [21]. These attributes reportedly influenced nutrient diffusion in the gel matrix [22]. Phytagel was reported to have a negative effect on AMF cultivation because of the impurities that hinder optimum growth and therefore there is a need of an alternative gelling agent [23]. On the other hand, pH of the culture also plays a significant role in AMF propagation, with optimal pH requirements varying with different species [24]. The pH of the growth medium in vitro can be influenced by various factors, including pre-autoclaving treatment, buffer additions, post-autoclaving adjustments, incubation duration, and light/dark conditions [25]. Live tissue cultures can also impact the medium's pH as they selectively absorb nutrients and release organic exudates [26]. The study highlighted the crucial role of Gellan gum's in supporting AMF growth and nutrient uptake while minimizing contamination, resulting in higher yields and better propagule quality. Its cost-effectiveness, attributed to the small amount needed for gelling, makes it a desirable choice for large-scale AMF mass production, given its superior performance in effective pH management.
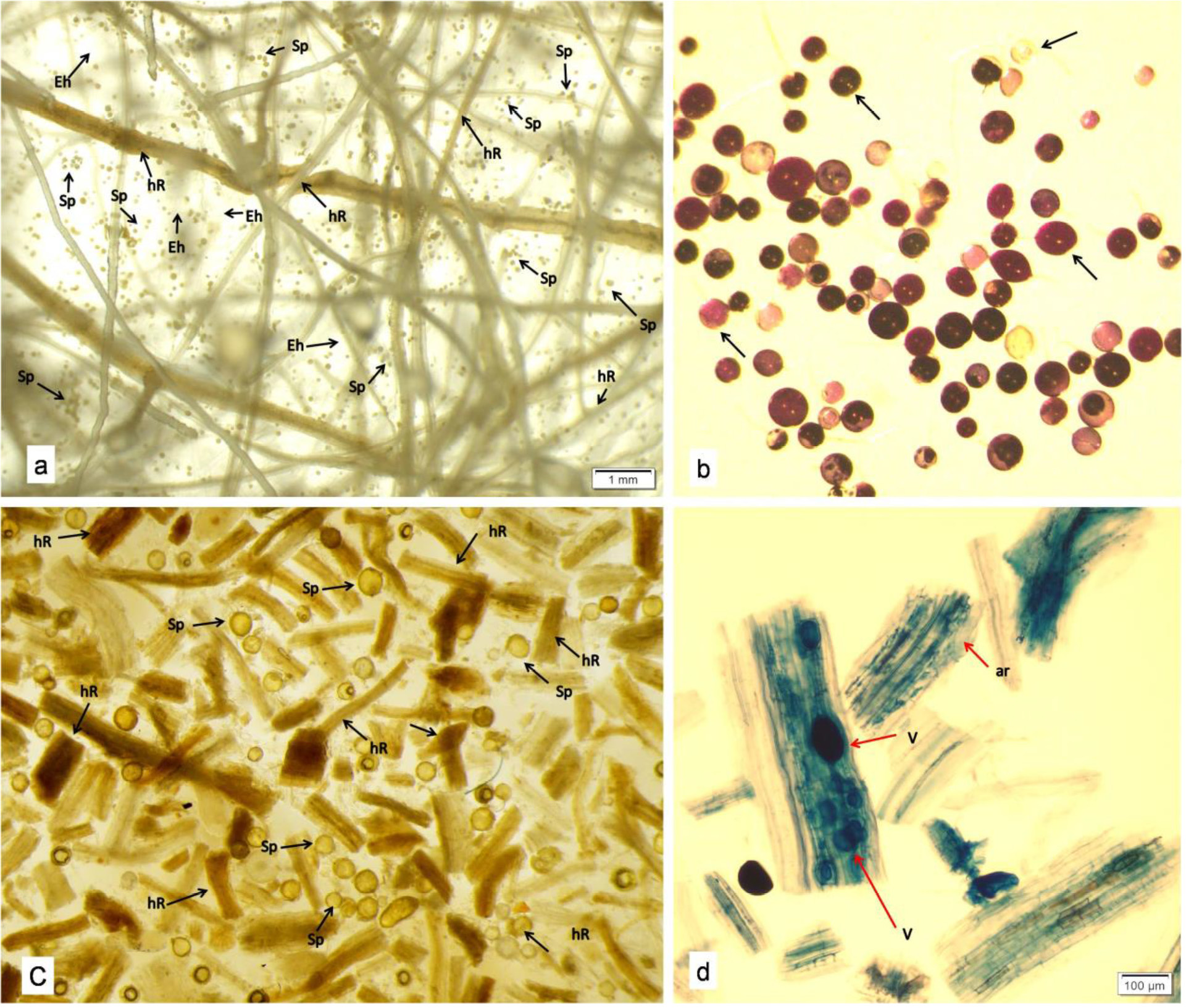
Figure 2 Final stages of in vitro AMF culturing in the current investigation. (a) Extraradical AMF spores observed in the ROC after 85 days of incubation showing healthy culture growth (Sp-Spores, hR-Host Roots, Eh-Extraradical hyphae) (b) Assessment of spore viability in end-product (c) The end-product, obtained through harvesting and subsequent drying, displayed AMF propagules, which included spores and roots fragments. (d) Presence of intraradical colonization of AMF inside the root fragments of the end-product verified their viability as propagules (V-Vesicles, ar-Arbuscules).
4.1.2. Media preparator
To achieve efficient and high-quality large-scale production, selecting equipment or approaches that maximize productivity is crucial. In our present investigation, we used media preparator for large scale production of culture media for AMF. The media preparator allowed for an automated operation with shorter processing time and proved to be more efficient and advanced. This study further indicated that, despite the higher cost, using a media preparator is more favorable than an autoclave due to its reduced contamination risk and improved end-product integrity. We therefore recommend the use of media preparator for large-scale production of culture medium.
4.2. Inoculation
4.2.1. Medium-to-inoculum ratio or starter culture volume
One of the important factors that influence culture quality and yield is the starter culture volume. There has been limited research on the optimal volume of starter culture for sub-culturing microorganisms and its impact on sporulation yield. Previous studies have shown that there is a correlation between the inoculum concentration and spore yield. A study conducted with Rhizophagus irregularis showed that the inoculum concentration of 0.6 DW g L-1 of medium yielded 20,000 spores L-1 [26]. Similarly, when 1 g of fresh weight (equivalent to 0.065 g DW) of inoculum was used, the final spore yield was 48,300 spores L-1 [27]. When 9.8 g fresh weight of VAM Rhizophagus irregularis was used as inoculum, the final spore yield was 14,300 spores L-1 in bioreactor over a period of 9 weeks while 3.7 g fresh weight of Rhizophagus aggregatus yielded 120,000 spores L-1 in bioreactor over a time of 8 weeks with reduced root growth in solid systems [28]. Another study documented the generation of 300,000 spores L-1 while utilizing a starter culture of 0.1-0.3 FW [29]. In our current investigation, a starter culture volume of 0.13 DW g L-1 was found to be optimal for achieving a remarkable average spore yield of 364,678 spores L-1 of medium consistently. This output was notably higher, being 10 to 18 times greater than the figures priorly reported by other researchers. Our large-scale production method demonstrated significant improvements in sporulation, leading to a substantial increase in spore production. To optimize large-scale production, determining the minimum starter culture volume that yields the best result is essential. Therefore, we suggest using a 0.13 dry weight inoculum for inoculating one litre of media, as this approach guarantees efficiency and cost-effectiveness for commercial sub-culturing.
4.2.2. Optimal relative humidity (RH) for culture incubation
While direct studies on the impact of RH on AMF in transformed root cultures are limited, it is widely acknowledged that maintaining optimal humidity levels is vital for successful in vitro cultivation of AMF [30]. RH regulates spore germination, and atmospheric moisture influences AMF proliferation and sporulation in their natural habitat, underscoring the importance of controlling RH for promoting their growth in vitro [30]. 25oC was cited as the optimal temperature for the long-term development of co-culture [8,31,32]. In our current study, we found that maintaining 50% RH at 25±2°C significantly reduced contamination levels, with a notable decrease in contamination rates from 32% (at ~60% RH) to 10% (at ~50% RH). These findings emphasize the importance of RH control for successful large-scale AMF cultivation.
4.2.3. Optimal culturing age with enhanced AMF yield
The age of the AMF culture is a crucial factor impacting spore and propagule yield, and this is supported by prior studies spanning 6 to 16 weeks of incubation. Spore production initially showed gradual growth up to 27 days of cultivation, followed by a significant increase up to day 76 when it stabilized [33]. Vegetative spores were abundant after 60 days of in vitro ROC of Rhizophagus irregularis. After 70 days, sporulation rates increased, possibly due to reduced sucrose levels in the medium [34]. Rhizophagus irregularis exhibited faster growth in vitro when placed in a sucrose-free medium. [35] Lower sucrose concentrations have been consistently associated with higher spore germination rates and increased sporulation [36]. Cultures maintained for 4-8 weeks, 8 weeks, 9 weeks, and 10-13 weeks yielded 2,766; 48,300; 120,000, and 30,000 spores L-1, respectively [37,27,28,26]. Extending the culture time to 12 weeks or beyond yielded significant results, with 280,000 spores L-1 [38] and up to 300,000 spores L-1 [34] within 12 weeks, 300,000 spores L-1 and 106 to 107 AMF propagules L-1 (sized >30 µ) in 6-12 weeks using the ROL technique [29] and 500,000 spores L-1 in 16 weeks [39]. In another study, incubation at 26±2ºC for 90 days using ROC with 0.4% CleriGel yielded 210,770±20,963 spores L-1 and 992,040±14,381 propagules spores L-1. Comparatively, M-media with 1% sucrose produced 556,940 spores L-1, while sugar-free MSR media yielded 196,390±7,524 spores L-1[40]. One methodology proposed for spore production achieved an average of 65,000 spores per plate in a 9-month process, involving manual root trimming and changing the entire medium every 2 months [41]. In contrast, another approach used two modified culture methods with cellophane and PVDF, producing nearly half the spore count per plate in half the time and without additional steps [42]. However, these methods were specifically designed for spore production, were time-consuming, labor-intensive, and carried contamination risks. In one study, approximately 60,250 spores were obtained per jar when using Solanum tuberosum, and over half of the roots were colonized within 12 weeks of inoculation [43]. Our study maintained a 12-week culture (particularly 85-88 days), resulting in about 2*106 AMF propagules L-1 of size >63 µ, indicating a substantial boost in propagule production. Thus, we recommend 12-week duration as the optimal culture period for large-scale AMF propagule production with Rhizophagus spp. in Carrot root ROCs.
4.3. Harvesting
4.3.1. Equipment
So far, no specific instrument has been designated for AMF harvesting, and there is no additional evidence suggesting that the choice of harvesting instrument influences the final product or yield. In one study, a food processor was used to shear roots from an aeroponic system, not for overall culture deionization [11]. In our current study, we compared two methods, namely, an orbital shaker and a blender (working like food processor), for deionization purposes. We observed a noteworthy disparity between the two methods. The liquid blender approach proved to be more effective and cost-efficient than the orbital shaker, especially in terms of root biomass yield and propagule yield, while there was no significant difference in spore yield and viability. Considering that large-scale production requires higher yields, we recommend employing the Blender Method for culture harvesting and large-scale deionization of AMF ROCs.
4.3.2. Optimal drying temperature for drying root biomass
While limited research was conducted on the influence of ideal drying temperature on the root biomass, only a single study suggests air-drying approach but did not provide precise temperature and the yield resulting from this method [7]. In our current investigation, drying the harvested roots at 25°C resulted in higher spore yield but necessitated longer drying time. Therefore, finding the right balance between maximizing spore production and optimizing process efficiency is essential when determining the suitable temperature.
Table 5. Comparison of different methodologies published earlier and the current investigation.
| Study | Technique | Gelling agent/Substrate | Media | Inoculum (Starter Culture) | Genus | Host | Incubation conditions | Duration | Spore, Propagule Yield | Biomass | Colonization |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Wood et al. [37]* | Plant ROC of herbaceous plants | Porous substrate (vermiculite & vermiculite-peat) and a nutrient solution | Not mentioned | 10-20 surface-sterilized pregerminated Gigaspora margarita spores | Rhizophagus, Gigaspora | Trifolium incarnatum, Arachis hypogaea | 25oC-28oC | 4-8 weeks | 2766 sp. L-1 | Up to 32% | |
| St-Arnaud et al. [39]* | Classical ROC | 0.5% w/v gellan gum | Modified M-medium | Not mentioned | Not mentioned | Daucus carota | 27oC | 4 months (16 weeks) | 500,000 sp. L-1 | Not mentioned | Not mentioned |
| Fortin et al. [28]* | Petri dish culture, Bioreactor | Agar (1%w/v), vermiculite | Modified White medium, MS medium, M-medium (pH 5.2 to 5.5) | 9.8 g FW 3.7g FW | Rhizophagus, Gigaspora, Scutellospora, Sclerocystis | Daucus carota, Trifolium pratense, etc | 24oC | 9 weeks | 48,300 sp. L-1 120,000 sp. L-1 | Not mentioned | Not mentioned |
| Jolicoeur et al. [26]* | Airlift bioreactor, Petri dish | Not mentioned | Low salt M-medium | 0.6 g DW g L-1 | Rhizophagus irregularis | Daucus carota | Not mentioned | 10-13 weeks | 20,000 sp. L-1 30,000 sp. L-1 | 0.13 g DW L-1 | 25%-75% |
| Declerck et al. [38]* | ROC in solid medium | Gel Gro (0.4% w/v) | MSR medium | 5 mm mycorrhizal root piece or spore | Rhizophagus | Daucus carota | Not mentioned | 12 weeks | 280,000 sp. L-1 | Not mentioned | Not mentioned |
| Wang [27]* | Not mentioned | Not mentioned | Not mentioned | 1 g FW (0.065 g DW) | Rhizophagus irregularis | Not mentioned | 26oC | 2 months (8 weeks) | 48300 sp. L-1 | 20.7± 0.57 | Not mentioned |
| Adholeya [7]* | Not mentioned (appears to be ROC) | Not mentioned | Not mentioned | Not mentioned | Rhizophagus, Gigaspora, Scutellospora | Not mentioned | Not mentioned | Not mentioned | Not mentioned | Not mentioned | Not mentioned |
| Puri & Adholeya [43]* | M-medium | Rhizophagus irregularis | Solanum tuberosum var ‘Pukhraj’ | 12 weeks | 60,250 spores/jar | ~400,000 sp. L-1 | >50% | ||||
| Srinivasan et al. [34]* | ROC in solid medium | Gallon gum (0.3% w/v) | MSR medium (pH 5.5 before autoclave) | 10-15 sterilized spores | Rhizophagus irregularis | Daucus carota | 27oC | 3 months | 300,000 sp. L-1 | Not mentioned | Not mentioned |
| Raj et al. [40]* | ROC with solid medium | 0.4% CleriGel (Phytagel in India) | Sugar free MW-media | Not mentioned | Rhizophagus irregularis | Daucus carota | 26±2ºC | 90 days | 210,770 sp. L-1; 992,040 ppg L-1 | Not mentioned | 75% |
| M-media fortified with 1% sugar | 556,940 sp. L-1 | ||||||||||
| Sugar free MSR media | 196,390sp. L-1 | ||||||||||
| Schuessler [29]* | Root organ liquid (ROL)-based ROC | Any group of polysaccharides like gellan gum, agar-agar, etc (0.1% or 0.05%w/v) | M-medium or MSR medium | 0.1-0.3 FW of AMF colonized root material | Acaulospora, Rhizophagus, Scutellospora, Funneliformis | Chicory, clover, bindweed, etc | 27oC | 42-84 days | 106 ppg L-1, 300,000 sp. L-1 | 80-120 mg FW L-1 | >53% |
| Current Study** | ROC in solid medium | Gellan gum (0.23% w/v) | M-medium (pH 5.5 before autoclave) | 0.13 g DW L-1 | Rhizophagus spp. | Daucus carota | 25oC, RH~50% | 85-88 days | >2*106 ppg L-1, >350,000 sp. L-1 | >2 g DW L-1 | 60-70% |
*The results incorporated in this tables from other studies are based on the published data
**The results incorporated in this table pertaining to the current study, are based on our experimental results and observations
**The results incorporated in this table pertaining to the current study, are based on our experimental results and observations
5. CONCLUSION
Our current research introduces a novel approach by customizing conventional ROC techniques for the purpose of large-scale AMF production. Our rigorous optimization efforts at every single stage of this process, using carrot root ROCs, led to substantial enhancements in both the quantity and quality of the final product, surpassing the results observed in earlier studies. This breakthrough underscores the effectiveness of our modifications, marking a significant advancement in the field of AMF production methods.
The optimized production processes offer several key benefits, including reduced contamination risk, improved control of nutrient diffusion, efficient pH management, higher yields, and improved propagule quality. The optimization of essential processes has notably influenced production costs, rendering it a more cost-efficient approach. This represents a promising method for large-scale AMF cultivation, yielding superior quality propagules.
Considering the increasing population, limited arable land and global demand for AMF inoculum, the production of high-quality AMF inoculum holds substantial economic potential. The standardized protocol outlined in our current study will greatly influence commercial-scale AMF production. These standardized protocols are poised to contribute to large-scale AMF production and carries significant implications for AMF applications in diverse agricultural and environmental contexts, with the potential to support sustainable agriculture and ecosystem management.
ACKNOWLEDGEMENTS
- Mycorrhizal symbiosis. Elsevier eBooks. 2008. https://doi.org/10.1016/b978-0-12-370526-6.x5001-6
- Biermann BJ, Linderman RG. Use of vesicular-arbuscular mycorrhizal roots, intraradical vesicles and extraradical vesicles as inoculum. New Phy. 1983 Sep;95(1):97–105. https://doi.org/10.1111/j.1469-8137.1983.tb03472.x
- Ghorui M, Chowdhury S, Balu P, Krishnan K, Djearamanec S, Manjunathana J, Preethi N, Ashokkumar KM, Jayanthi M. A review: in vitro cultivation of Arbuscular Mycorrhizal Fungus for commercialization. Oxi Com. 2023;46(3):549–564.
- Basiru S, Mwanza HP, Hijri M. Analysis of Arbuscular mycorrhizal fungal inoculant benchmarks. Microorganisms. 2020 Dec;9(1):81. https://doi.org/10.3390/microorganisms9010081
- Adholeya A, Tiwari P, Singh R. Large-Scale inoculum production of arbuscular mycorrhizal fungi on root organs and inoculation strategies. Soil bio. 2005. p. 315–338. https://doi.org/10.1007/3-540-27331-x_17
- Goh D, Martin JGA, Banchini C, MacLean AM, Stefani F. RocTest: A standardized method to assess the performance of root organ cultures in the propagation of arbuscular mycorrhizal fungi. Fro. in Micro. 2022 Jul 28;13. https://doi.org/10.3389/fmicb.2022.937912
- Alok A. WO2013098829A1 - Novel mycorrhizae-based biofertilizer compositions and method for mass production and formulations of same - Google Patents. 2011. https://patents.google.com/patent/WO2013098829A1/en
- Bécard G, Fortin JA. Early events of vesicular–arbuscular mycorrhiza formation on Ri T‐DNA transformed roots. New Phytologist. Wiley-Blackwell; 1988 Feb;108(2):211–218. https://doi.org/10.1111/j.1469-8137.1988.tb03698.x
- Sandle T PhD. Assessment of Culture Media in Pharmaceutical Microbiology. https://www.americanpharmaceuticalreview.com/Featured-Articles/163589-Assessment-of-Culture-Media-in-Pharmaceutical-Microbiology/
- Doner LW, Bécard G. Solubilization of gellan gels by chelation of cations. Biotech. Tech.; 1991 Jan;5(1):25–28. https://doi.org/10.1007/bf00152749
- Sylvia DM, Jarstfer AG. Sheared-Root inocula of Vesicular-Arbuscular mycorrhizal fungi. Applied and Environmental Microbiology. Ame. Soc. for Micro.; 1992 Jan;58(1):229–232. https://doi.org/10.1128/aem.58.1.229-232.1992
- An Z, Bin G, Hendrix JW. Viability of soilborne spores of glomalean mycorrhizal fungi. Soil Biol. & Biochem.; 1998 Aug;30(8–9):1133–1136. https://doi.org/10.1016/s0038-0717(97)00194-6
- Phillips J, Hayman DS. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. of the Bri. Myc. Society; 1970 Aug;55(1):158-IN18. https://doi.org/10.1016/s0007-1536(70)80110-3
- Cell Culture Contamination | Thermo Fisher Scientific - IE. https://www.thermofisher.com/in/en/home/references/gibco-cell-culture-basics/biological-contamination.html
- R: The R Project for Statistical Computing. https://www.r-project.org/
- Ellatif SA, Ali E, Senousy HH, Razik ESA. Production of Arbuscular Mycorrhizal Fungi using In vitro Root Organ Culture and Phenolic Compounds. J. of Pure and App. Micro. 2019 Dec;13(4):1985–94. https://doi.org/10.22207/jpam.13.4.10
- Das N, Tripathi N, Basu S, Bose C, Maitra S, Khurana S. Progress in the development of gelling agents for improved culturability of microorganisms. Frontiers in Microbiology. 2015 Jul;6. https://doi.org/10.3389/fmicb.2015.00698
- Jaramillo J, Summers WL. Tomato anther callus production: Solidifing agent and concentration influence induction of callus. J. of the Ame. Soc. for Hor. Sci. 1990 Nov;115(6):1047–50. https://doi.org/10.21273/jashs.115.6.1047
- McGuffey JC, Leon D, Dhanji EZ, Mishler DM, Barrick JE. Bacterial production of gellan gum as a Do-It-Yourself alternative to Agar. J. of Micro. & Bio. Edu. 2018 Jan;19(2). https://doi.org/10.1128/jmbe.v19i2.1530
- All you need to know about Gellan Gum. Plant Cell Technology | Your Partner in Plant Tissue Culture. https://www.plantcelltechnology.com/blogall-you-need-to-know-about-gellangum/#:~:text=The%20gellan%20gum%20has%20more,plant%20tissue%20prior%20to%20transplanting
- Mao R, Tang J, Swanson BG. Water holding capacity and microstructure of gellan gels. Carb. Pol. 2001 Dec;46(4):365–71. https://doi.org/10.1016/s0144-8617(00)00337-4
- Gupta SD, Ibaraki Y. Plant tissue Culture Engineering. Springer Science & Business Media; 2006.
- Bhatia NP, Adholeya A, Verma A. Influence of media gelling agents on root biomass and in vitro VA-mycorrhizal symbiosis of carrot with Gigaspora margarita. Biotropia: The Sou. Asi. J. of Tro. Bio. 1997 Jun;0(10). https://doi.org/10.11598/btb.1997.0.10.128
- Tiwari P, Adholeya A. In vitro co-culture of two AMF isolates Gigaspora margarita and Glomus intraradices on Ri T-DNA transformed roots. Fems Micro. Let. 2002 Jan;206(1):39–43. https://doi.org/10.1111/j.1574-6968.2002.tb10983.x
- Owen H, Wengerd D, Miller A. Culture medium pH is influenced by basal medium, carbohydrate source, gelling agent, activated charcoal, and medium storage method. Plant Cell Rep. 1991 Dec;10(11). https://doi.org/10.1007/bf00232516
- Jolicœur M, Williams RD, Chavarie C, Fortin JA, Archambault J. Production of Glomus intraradices propagules, an arbuscular mycorrhizal fungus, in an airlift bioreactor. Biotechnology and Bioengineering. 1999 Apr;63(2):224–32. https://doi.org/10.1002/(sici)1097-0290(19990420)63:2<224::aid-bit11>3.0.co;2-x
- Wang WK. US6759232B2 - Method of facilitating mass production and sporulation of arbuscular mycorrhizal fungi aseptic in vitro - Google Patents. 2002. https://patents.google.com/patent/US6759232B2/en
- Fortin JA. US5554530A - Aseptic in vitro endomycorrhizal spore mass production - Google Patents. 1993. https://patents.google.com/patent/US5554530A/en
- Schuessler A. EP3038456A1 - System and methods for continuous propagation and mass production of arbuscular mycorrhizal fungi in liquid culture - Google Patents. 2013. https://patents.google.com/patent/EP3038456A1/en
- Siqueira JO, Sylvia DM, Gibson J, Hubbell DH. Spores, germination, and germ tubes of vesicular–arbuscular mycorrhizal fungi. Can. J. of Micro. 1985 Nov;31(11):965–72. https://doi.org/10.1139/m85-183
- Engelmoer DJP, Behm JE, Kiers ET. Intense competition between arbuscular mycorrhizal mutualists in an in vitro root microbiome negatively affects total fungal abundance. Mol. Eco. 2013 Sep;23(6):1584–93. https://doi.org/10.1111/mec.12451
- Danesh YR, Goltapeh EM, Alizadeh AHM, M. Modarres Sanavy. Optimizing carrot hairy root production for monoxenic culture of arbuscular mycorrhizal fungi in Iran. J. of Bio. Sci. 2005 Dec. https://doi.org/10.3923/jbs.2006.87.91
- Costa FA, Haddad LSM, Kasuya MCM, Oton WC, Costa MD, Borges AC. In vitro culture of Gigaspora decipiens and Glomus clarum in transformed roots of carrot: the influence of temperature and pH. Acta Scientiarum-agronomy. 2013 Jul;35(3). https://doi.org/10.4025/actasciagron.v35i3.16581
- Srinivasan M, Kumar KK, Kumutha K, Marimuthu P. Establishing monoxenic culture of arbuscular mycorrhizal fungus Glomus intraradices through root organ culture. J. of App. and Nat. Sci. 2014 Jun 1;6(1):290–3. https://doi.org/10.31018/jans.v6i1.417
- D’Souza J. Modified Strullu and Romand (MSR) medium devoid of sucrose promotes higher in vitro germination in Rhizophagus irregularis. J. of Myc. and Plant Path., 43(2), 240–24. 2013. http://irgu.unigoa.ac.in/drs/handle/unigoa/2853
- Dalpé Y, De Souza FA, Declerck S. Life cycle of glomus species in monoxenic culture. In: Soil biology. 2005. p. 49–71. https://doi.org/10.1007/3-540-27331-x_4
- Wood T. EP0209627A2 - Method for producing axenic vesicular arbuscular mycorrhizal fungi in association with root organ cultures - Google Patents. 1985. https://patents.google.com/patent/EP0209627A2/en
- Declerck S, D’Or D, Cranenbrouck S, Boulengé LE. Modelling the sporulation dynamics of arbuscular mycorrhizal fungi in monoxenic culture. Mycorrhiza. 2001 Oct;11(5):225–30. https://doi.org/10.1007/s005720100124
- St‐Arnaud M, Hamel C, Vimard B, Caron M, Fortin JA. Enhanced hyphal growth and spore production of the arbuscular mycorrhizal fungus Glomus intraradices in an in vitro system in the absence of host roots. Myc. Res. 1996 Mar;100(3):328–32. https://doi.org/10.1016/s0953-7562(96)80164-x
- Raj BM, Kumar RB, Rao GV, Murthy KSR. An optimised in vitro protocol for mass production of Rhizophagus irregularis spores - for sustainable agriculture. J. of Bus. Res. 2017 Jul;9(4):21–9. https://africaneditors.org/journal/JBR/full-text-pdf/66716-106667
- Douds DD. Increased spore production by Glomus intraradices in the split-plate monoxenic culture system by repeated harvest, gel replacement, and resupply of glucose to the mycorrhiza. Mycorrhiza. 2002 Aug;12(4):163–7. https://doi.org/10.1007/s00572-002-0174-9
- Rosikiewicz P, Bonvin JF, Sanders IR. Cost-efficient production of in vitro Rhizophagus irregularis. Mycorrhiza. 2017 Feb;27(5):477–86. https://doi.org/10.1007/s00572-017-0763-2
- Puri A, Adholeya A. A new system using Solanum tuberosum for the co-cultivation of Glomus intraradices and its potential for mass producing spores of arbuscular mycorrhizal fungi. Symbiosis. 2013 Jan;59(2):87–97. https://doi.org/10.1007/s13199-012-0213-z
The authors are grateful to Symbiotic Sciences Pvt Ltd, India for the support extended during this study.