
Download


Journal of Biological Methods | 2023 | Vol. 10 | e99010004
DOI: 10.14440/jbm.2023.403
DOI: 10.14440/jbm.2023.403
PROTOCOL
An experimental workflow for identifying RNA m6A alterations in cellular senescence by methylated RNA immunoprecipitation sequencing
Yue Shi2,3,†, Zeming Wu1,4,†, Weiqi Zhang2,3,5, Jing Qu4,5,6, Weimin Ci2,3,5,*, Guang-Hui Liu1,4,5,*
1State Key Laboratory of Membrane Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China
2CAS Key Laboratory of Genomic and Precision Medicine, Beijing Institute of Genomics, Chinese Academy of Sciences, Beijing 100101, China
3China National Center for Bioinformation, Beijing 100101, China
4Beijing Institute for Stem Cell and Regenerative Medicine, Beijing 100101, China
5University of Chinese Academy of Sciences, Beijing 100049, China
6State Key Laboratory of Stem Cell and Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China
2CAS Key Laboratory of Genomic and Precision Medicine, Beijing Institute of Genomics, Chinese Academy of Sciences, Beijing 100101, China
3China National Center for Bioinformation, Beijing 100101, China
4Beijing Institute for Stem Cell and Regenerative Medicine, Beijing 100101, China
5University of Chinese Academy of Sciences, Beijing 100049, China
6State Key Laboratory of Stem Cell and Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China
†These authors contributed equally to this work.
*Corresponding author: Guang-Hui Liu Tel: +86 10 64807852; Email: ghliu@ioz.ac.cn; ORCID: 0000-0001-9289-8177; Weimin Ci Tel: +86 10 84097318; Email: ciwm@big.ac.cn; ORCID: 0000-0002-4111-5360.
Competing interests: The authors have declared that no competing interests exist.
Abbreviation used: m6A, N6-methyladenosine; meRIP-seq, methylated RNA immunoprecipitation followed by high-throughput sequencing; RT-qPCR, reverse-transcription quantitative PCR; RT, room temperature; SA-β-gal, senescence-associated beta-galactosidase.
Received April 27, 2023; Revision received July 6, 2023; Accepted July 7, 2023; Published August 25, 2023
Abstract
N6-methyladenosine (m6A), the most prevalent mRNA modification in eukaryotic cells, is known to play regulatory roles in a wide array of biological processes, including aging and cellular senescence. To investigate such roles, the m6A modification can be identified across the entire transcriptome by immunoprecipitation of methylated RNA with an anti-m6A antibody, followed by high-throughput sequencing (meRIP-seq or m6A-seq). Presented here is a protocol for employing meRIP-seq to profile the RNA m6A landscape in senescent human cells. We described, in detail, sample preparation, mRNA isolation, immunoprecipitation, library preparation, sequencing, bioinformatic analysis and validation. We also provided tips and considerations for the optimization and interpretation of the results. Our protocol serves as a methodological resource for investigating transcriptomic m6A alterations in cellular senescence as well as a valuable paradigm for the validation of genes of interest.
Keywords: m6A, cellular senescence, meRIP-seq, bioinformatic analysis, qPCR
INTRODUCTION
Cellular senescence represents an intricate biological process characterized by irreversible growth arrest and changes in gene expression patterns [1-4]. Senescence is also as a hallmark of organismal aging and a risk factor for a variety of age-related diseases [2,5-8]. Recent studies have demonstrated that epi-transcriptomic modifications, including N6-methyladenosine (m6A), are involved in the regulation of cellular senescence [9,10]. As the most common mRNA modification in eukaryotic cells, m6A impacts splicing, stability, and translation of RNA, playing a key role in the fine-tuning of gene expression associated with senescence regulation [11-15]. For example, m6A modifications reportedly affected the stability and translation of key downstream mRNAs, thereby accelerating or impeding the progression of cellular senescence [14-16]. Accordingly, deciphering the role and mechanism of m6A in cellular senescence can help identify novel biomarkers for aging and potential targets for the management of aging-associated disorders.
Methylated RNA immunoprecipitation plus ensuing high-throughput sequencing (meRIP-seq or m6A-seq) has been commonly used for the identification of m6A modifications in RNA [17-20]. With this technique, m6A-containing RNAs are immunoprecipitated by using an anti-m6A antibody and then subjected to high-throughput sequencing. This approach allows for the transcriptome-wide identification of m6A sites, providing invaluable insights into the role of m6A in cellular processes. Several studies have applied meRIP-seq to investigate the changes of m6A in cellular senescence. For instance, two representative studies found that m6A modifications were decreased in senescent cells compared to non-senescent cells and identified novel m6A-regulated genes implicated in senescence [14,21]. Despite its broad application, several considerations should be taken when using meRIP-seq to study m6A modifications in cellular senescence. One crucial factor is sample preparation, since acquiring high-quality RNA is crucial to the success of the meRIP-seq. Antibody selection is another important consideration, as specificity can affect the accuracy of the immunoprecipitation step. In addition, sequencing depth can influence the sensitivity of the results.
In this study, we worked out a workflow for using meRIP-seq to study RNA m6A modifications in control (non-senescent) and senescent human cells, which included operational steps for sample preparation, mRNA isolation, immunoprecipitation, library preparation, sequencing, bioinformatic analysis and target gene validation via reverse-transcription quantitative PCR (RT-qPCR). We also provided tips and considerations in the optimization of the protocol and interpretation of the results. Overall, this workflow provided a detailed procedure for investigating transcriptomic m6A alterations and identifying downstream targets of interest in cellular senescence.
MeRIP-seq & meRIP-qPCR protocols
MATERIALS
Chemicals/reagents/others
- TRIzol™ Reagent (Cat. #15596026, Invitrogen, Thermo Fisher Scientific, Shanghai, China)
- Chloroform (Cat. #C2432, Sigma, Shanghai, China)
- RNase-free EP tube (Cat. #MCT-150-C, Axygen, Shanghai, China)
- Isopropanol (Cat. #AC184130010, Invitrogen, Thermo Fisher Scientific)
- Ethanol (Cat. #18-606-046, Invitrogen, Thermo Fisher Scientific)
- Qubit™ RNA HS Assay (Cat. #Q32852, Invitrogen, Thermo Fisher Scientific)
- Qubit™ RNA XR Assay (Cat. #Q33223, Invitrogen, Thermo Fisher Scientific)
- Qubit™ RNA IQ Assay Kits (Cat. #Q33221, Invitrogen, Thermo Fisher Scientific)
- Anti-m6A antibody (Cat. #202003, Synaptic Systems, Goettingen, Germany)
- Dynabeads™ Protein A (Cat. #10001D, Invitrogen, Thermo Fisher Scientific)
- Nuclease-free water (Cat. #AM9937, Invitrogen, Thermo Fisher Scientific)
- Dynabeads™ mRNA Purification Kit (Cat. #61006, Invitrogen, Thermo Fisher Scientific)
- DNase I (Cat. #M0303, NEB, Beijing, China)
- RNA Fragmentation Reagents (Cat. #AM8740, Invitrogen, Thermo Fisher Scientific)
- N6-methyladenosine (Cat. #1867-73-8, Berry & Associates, Bishop Circle East Dexter, MI)
- RNasin® Ribonuclease Inhibitor (Cat. #N2515, Promega, Beijing, China)
- Acid Phenol: Chloroform (pH 4.3-4.7) (Cat. #AM9720, Invitrogen, Thermo Fisher Scientific)
- Glycogen (20 mg/mL) (Cat. #R0551, Invitrogen, Thermo Fisher Scientific)
- 3 M Sodium Acetate, pH 5.5 (Cat. #AM9740, Invitrogen, Thermo Fisher Scientific)
- 1 M Tris-HCl (pH 7.4) (Cat. #AM9850, Invitrogen, Thermo Fisher Scientific)
- 5 M NaCl (Cat. #AM9759, Invitrogen, Thermo Fisher Scientific)
- 10% NP-40 (Cat. #85124, Invitrogen, Thermo Fisher Scientific): 10% (w/v) solution in water.
- DEPC-treated water (Cat. #B501005-0500, Sangon, Shanghai, China)
- PBS (Cat. #10010023, Invitrogen, Thermo Fisher Scientific)
- Stranded RNA-Seq Library Preparation Kit (Cat. #KK8401, KAPA Biosystems, Roche, Wilmington, MA)
- GoScript™ Reverse Transcription System (Cat. #A5001, Promega)
- THUNDERBIRDTM SYBR® qPCR mix (Cat. #QPS-201, TOYOBO, Osaka, Japan)
- IP buffer:
| Materials | Volume |
|---|---|
| Tris-HCl (1 M, pH 7.4) | 1 mL |
| NaCl (5 M) | 3 mL |
| NP-40% (10%) | 1 mL |
| DEPC-treated water | 95 mL |
| Total | 100 mL |
- Low-salt IP buffer:
| Materials | Volume |
|---|---|
| Tris-HCl (1 M, pH 7.4) | 1 mL |
| NaCl (5 M) | 1 mL |
| NP-40% (10%) | 1 mL |
| DEPC-treated water | 97 mL |
| Total | 100 mL |
- High-salt IP buffer:
| Materials | Volume |
|---|---|
| Tris-HCl (1 M, pH 7.4) | 1 mL |
| NaCl (5 M) | 10 mL |
| NP-40% (10%) | 1 mL |
| DEPC-treated water | 88 mL |
| Total | 100 mL |
Equipment
- Centrifuge (Cat. #MicroCL 21R, Invitrogen, Thermo Fisher Scientific)
- Block heaters (Cat. #88870006, Invitrogen, Thermo Fisher Scientific)
- General rotator (Cat. #88882016, Invitrogen, Thermo Fisher Scientific)
- Qubit 4 Fluorometer (Cat. #Q33226, Invitrogen, Thermo Fisher Scientific)
- T100 Thermal Cycler (Cat. #1861096, Bio-Rad, Beijing, China)
- QuantStudio™ 5 Real-Time PCR System (Cat. #A28140, Applied Biosystems, Thermo Fisher Scientific)
Software packages
- MACS2 (Version 2.2.2)
- BEDTools (Version 2.25.0)
- HOMER (Version 4.11)
- IGV (Version 2.3.34)
PROCEDURES AND RESULTS
A schematic representation for the workflow of meRIP-seq experiment in young and senescent cells is shown in Fig. 1. The steps are detailed as follows:
1.Sample collection and total RNA extraction (Timing 3 hours)
1.1.When the cells are virtually confluent, remove the culture medium, wash the cells with 2‒5 mL PBS, and aspirate the PBS.
Tips: A senescence phenotype test, e.g., senescence-associated beta-galactosidase (SA-β-gal) staining, needs to be performed on the cells before collection.
1.2.Directly add 1 mL TRIzol™ Reagent to the culture dish (10 cm) to lyse 1 × 105-107 cells and lyse the cells by pipetting the TRIzol™ reagent up and down several times.
1.3.Transfer the lysate to RNase-free EP tubes and incubate the tubes at room temperature (RT) for 5 minutes.
1.4.Add 0.2 mL chloroform to the lysate, vortex the mixture for 15 seconds and incubate the mixture at RT for 5 minutes.
1.5.Centrifuge the lysate for 15 minutes at 12,000 ×g at 4°C.
1.6.Transfer the RNA-containing aqueous phase to a new RNase-free EP tube.
1.7.Add an equal volume of pre-cooled isopropanol and incubate the mixture for 10 minutes on ice.
1.8.Centrifuge the sample for 10 minutes at 12,000 ×g at 4°C.
1.9.Discard the supernatant and wash total RNA once with 1 mL 70% ethanol.
1.10.Centrifuge the tube at 14,800 rpm and 4℃ for 15 minutes. Discard the supernatant.
1.11.Allow the RNA to air dry for 10‒15 minutes. Dissolve the total RNA in 100‒200 μL nuclease-free water. Measure the total RNA concentration and quality on a Qubit 4 Fluorometer by using Qubit™ RNA XR Assay and Qubit™ RNA IQ Assay Kits.
Note: Isolated total RNA can be stored at -80 °C for up to 6 months until further use.
Figure 1. Workflow for meRIP-seq and validation in young and senescent cells. A schematic diagram showing the meRIP-seq procedures, including sample preparation, total RNA extraction and mRNA isolation, immunoprecipitation, sequencing and bioinformatic analysis, and meRIP-RT-qPCR validation for genes of interest during cellular senescence.
2.mRNA isolation (Timing 3 hours)
2.1.Add nuclease-free water to total RNA to adjust volume of 75 μg total RNA to 100 μL. Incubate the total RNA for 2 minutes at 65°C to disrupt secondary structures, then place the RNA on ice.
2.2.Transfer 200 μL (1 mg) of well-resuspended Dynabeads to a new 1.5 mL EP tube. Place the tube on the magnet and make sure all beads migrate to the magnet-touching wall of the tube.
2.3.Discard the supernatant and remove the tube from the magnet. Then add 100 μL Binding Buffer to the beads, place the tube back on the magnet to let all beads migrate to the magnet-touching wall of the tube, and then remove the supernatant.
2.4.Mix the Dynabeads with 100 μL Binding Buffer and 100 μL total RNA thoroughly, and rotate the mixture on a general rotator for 10 minutes at RT.
2.5.Place the tube on the magnet, let all beads migrate to the magnet-touching wall of the tube and then discard the supernatant.
2.6.Remove the tube from the magnet and add 200 μL Washing Buffer B to the mRNA-beads complex. Pipette the mixture up and down several times. Then place the tube on the magnet, allow all beads to migrate to the magnet-touching wall of the tube and then discard the supernatant. Repeat this step once more.
2.7.Add 20‒50 μL nuclease-free water to elute mRNA from the Dynabeads. Heat the samples at 70°C for 2 minutes. Then, place the tube on the magnet. Next, elute the mRNA and transfer it to a new RNase-free tube. Measure the mRNA concentration on the Qubit 4 Fluorometer with Qubit™ RNA HS Assay Kits.
3.DNase I digestion (Timing 1.5 hours)
3.1.Mix 5 μL 10× DNase I buffer and 2 μL DNase I with the purified mRNA to have a final volume at 50 μL. Then incubate the sample at 37℃ for 10 minutes to digest any remaining cellular DNA.
3.2.Add 2 μL glycogen, 5 μL 3 M NaOAc (pH 5.5), and 190 μL 100% ethanol to the mRNA-DNase I mixture to precipitate the mRNA. Incubate the sample for 1‒2 hours or overnight at -80℃.
3.3.Precipitate the mRNA from the last step by centrifuging at 14,800 rpm and 4℃ for 40‒50 minutes. Wash the mRNA once with 1 mL 70% ethanol. Centrifuge the tube at 14,800 rpm and 4℃ for 15 minutes. Discard the supernatant.
Tips: It is desirable to centrifuge again after discarding the 100% ethanol to remove the remaining ethanol and dry the mRNA completely, since ethanol carryover into the eluate may interfere with some downstream procedures.
3.4.Dry the mRNA at RT for 10 minutes. Resolve the mRNA with 20 μL nuclease-free water. Measure the mRNA concentration on the Qubit 4 Fluorometer by using Qubit™ RNA HS Assay Kits.
3.5.Separate 100‒200 ng mRNA as input for RNA-seq.
4.RNA fragmentation and purification (Timing 1.5 hours)
4.1.Add 1 μL of RNA Fragmentation Reagent into each 10 μL mRNA in PCR tube, and incubate the sample for 50 seconds at 90°C to obtain 100‒200 nt fragments (no more than 2 μg/tube).
4.2.Pool all the mRNA together. Add 2 μL glycogen, 5 μL 3 M NaOAc (pH 5.5), and 190 μL 100% ethanol to the fragmented mRNA. Incubate the mixture at -80°C for 1‒2 hours or overnight.
4.3.Precipitate the fragmented mRNA from the last step by centrifuging at 14,800 rpm and 4℃ for 40‒50 minutes. Wash it once with 1 mL 70% ethanol. Centrifuge the sample at 14,800 rpm and 4℃ for 15 minutes. Discard the supernatant.
4.4.Dry the mRNA at RT for 10 minutes. Resolve the mRNA with 20 μL nuclease-free water. Measure the concentration of the mRNA on the Qubit 4 Fluorometer by using the Qubit™ RNA HS Assay Kits.
Tips: It is desirable to centrifuge the resultant sample again after discarding the 100% ethanol to remove the rest ethanol and dry the mRNA completely, since ethanol carryover into the eluate may interfere with some downstream procedures.
Note: Fragmented RNA can be stored at -80 °C for up to 6 months until further use.
5.m6A-tagged RNA pull down (immunoprecipitation), library construction and high-throughput sequencing (Timing 5 hours)
5.1.Transfer 20 μL beads (for each 2 μg mRNA and 5 μg anti-m6A antibody) to a new RNase-free tube.
5.2.Wash the beads three times in 1 mL 1× IP buffer for 3‒5 minutes with gentle rotation at RT. Place the tube on the magnet until all beads have migrated to the magnet-touching wall of the tube, and discard the supernatant.
5.3.Resuspend the beads in 500 μL 1× IP buffer containing 2 μL RNasin and 5 μg anti-m6A antibody and incubate the sample for 1 hour with gentle rotation at RT.
5.4.Wash the antibody-bead mixture three times with 1 mL 1× IP buffer for 3‒5 minutes with gentle rotation.
5.5.Incubate the fragmented mRNA for 2 minutes at 70°C to disrupt secondary structures, and then place it on ice.
5.6.Add 2 μg fragmented RNA to the antibody-beads mixture, and incubate the mixture for 4 hours with gentle rotation at 4ºC.
5.7.Wash the mRNA-antibody-bead mixture three times with 1 mL 1× IP buffer for 2 minutes with gentle rotation at RT.
5.8.Repeat last step with 1 mL low-salt buffer for 2 minutes with gentle rotation at RT.
5.9.Repeat last step with 1 mL 1× IP buffer for 2 minutes with gentle rotation at RT.
5.10.Repeat last step with 1 mL high-salt buffer for 2 minutes with gentle rotation at RT.
5.11.Repeat last step with 1 mL 1× IP buffer for 2 minutes with gentle rotation at RT.
5.12.Competitively elute the mRNA with m6A modification by adding 300 μL 0.5 mg/mL N6-methyladenosine and 3 μL RNasin in IP buffer and incubating the resultant sample with gentle rotation at RT for 1 hour.
5.13.Repeat last step once more.
5.14.Pool the eluates together, and add equal volume of Acid Phenol: Chloroform. Vortex the sample thoroughly, then incubate them on ice for 5 minutes.
5.15.Precipitate the eluted mRNA from the last step by centrifugation at 14,800 rpm and 4℃ for 15 minutes and transfer the supernatant to two new 1.5 mL EP tubes.
5.16.Add 2 μL glycogen, 30 μL 3 M NaOAc, and 3 volumes of 100% ethanol to precipitate the eluted mRNA by incubation at -80°C overnight.
5.17.Precipitate the mRNA from the last step by centrifugation at 14,800 rpm and 4℃ for 40‒50 minutes. Wash the mRNA once with 1 mL 70% ethanol. Centrifuge the tube at 14,800 rpm and 4℃ for 1 minute. Discard the supernatant.
5.18.Dry the mRNA at RT for 10 minutes. Dissolve the mRNA in 10 μL nuclease-free water. At this point, the samples can be used for subsequent library construction or RT-qPCR.
5.19.Use KAPA Stranded RNA-Seq Library Preparation Kits for library construction according to the manufacturer’s instructions.
5.20.Subject libraries to cluster generation and next-generation sequencing on an Illumina HiSeq X Ten platform.
Tips: In order to lower the risk of missing peaks as a result of suboptimal coverage, it is essential that the average gene coverage be within the range of 10–50×.
Note: The eluted RNA can be stored at -80 °C for up to 6 months until further use.
6.Bioinformatic analysis for meRIP-seq [14,15]
6.1.Use the MACS2 (Version 2.2.2) for m6A peak calling with the default options except for ‘–nomodel, –keepdup all’. A P-value cutoff of 1×10−5 is used for the high-confidence peaks.
6.2.Intersect each peak from Step 6.1 with the UCSC gene annotation bed file by using BEDTools ‘IntersectBed’ (Version 2.25.0) for peak annotation.
6.3.Use HOMER (Version 4.11) for motif enrichment analysis by setting a motif length of 6 nucleotides.
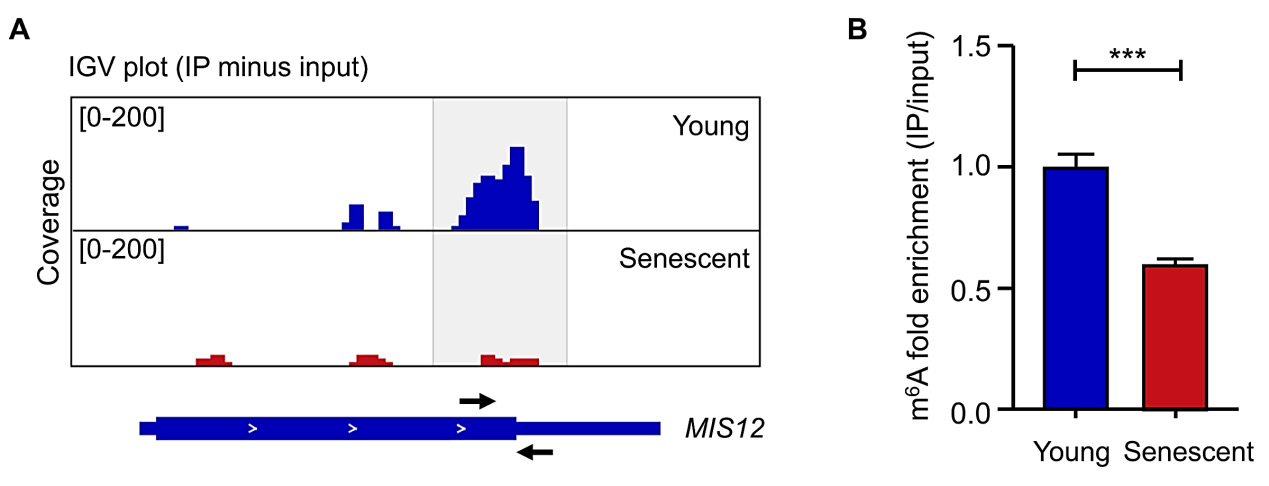
6.4.Generate the strand-specific bigwig files from RNA-seq (input) and meRIP-seq (IP) by BedTools (Version 2.25.0) and visualized by IGV (Version 2.3.34) (Fig. 2A).
7.MeRIP-qPCR validation
7.1.Primer design
Design primers targeting genes of interest in the transcript region where the m6A peaks are located based on the meRIP-seq results. Taking human MIS12 as an example [14], the binding sites for primers used for RT-qPCR validation are shown in Fig. 2A. Sequence information of primers targeting human MIS12 is listed below:
Human-MIS12-Fwd: 5'-TGTTGGCAGAGATCATGGGAC-3'
Human-MIS12-Rev: 5'-TGACAGGCTCCTCCTTTTGAC-3'
7.2.MeRIP
See details for mRNA isolation, DNase I digestion, RNA fragmentation and purification and immunoprecipitation in PROCEDURES AND RESULTS above. Equalize the final volume of input and IP RNA of control and senescent cells to 10 μL in nuclease-free water. The resulting input and IP RNA samples are used for subsequent RT-qPCR experiment.
7.3.RT-qPCR
7.3.1.Reverse transcription
Subject equal amounts of RNA from each sample to reverse transcription using a GoScript Reverse Transcription System by following the manufacturer's instructions. The procedure is described in detail as follows:
7.3.2.Prepare the following materials required for the first-step reaction and mix them thoroughly.
Components for the first-step reaction of reverse transcription:
| Materials | Volume |
|---|---|
| RNA | 5 μL |
| Random Primers | 1 μL |
| Oligo (dT)15 Primers | 1 μL |
| Nuclease-free water | 3 μL |
7.3.3.Briefly centrifuge the aforementioned mixture and start the first-step reaction of reverse transcription under the following conditions.
Conditions for the first-step reaction of reverse transcription:
| Steps | Temperature | Time |
|---|---|---|
| I | 70°C | 5 min |
| II | 4°C | ∞ |
7.3.4.Prepare the following components required for the second-step reaction and add them into the above-mentioned reaction mixture.
Components for the second-step reaction of reverse transcription:
| Materials | Volume |
|---|---|
| Nuclease-free water | 1.6 μL |
| 5×Reaction Buffer | 4 μL |
| 25 mM MgCl2 | 2 μL |
| PCR Nucleotide Mix | 1 μL |
| Reverse Transcriptase | 1 μL |
| Ribonuclease Inhibitor | 0.4 μL |
7.3.5.Thoroughly mix and briefly centrifuge the above mixture and start the second-step reaction of reverse transcription by using the following conditions.
Conditions for the second-step reaction of reverse transcription:
| Steps | Temperature | Time |
|---|---|---|
| I | 25°C | 5 min |
| II | 42°C | 60 min |
| III | 70°C | 15 min |
| IV | 4°C | ∞ |
7.3.6.Prepare qPCR samples as indicated below and mix them thoroughly.
qPCR mix:
| Materials | Volume |
|---|---|
| Nuclease-free water | 2 μL |
| cDNA | 0.1 μL |
| SYBR® qPCR Mix | 2.5 μL |
| Forward primer (10 μM) | 0.2 μL |
| Reverse primer (10 μM) | 0.2 μL |
7.3.7.Briefly centrifuge the above mixture and run qPCR using the following conditions.
qPCR conditions:
7.4.Calculate the fold enrichment of m6A (IP/input) on MIS12 mRNA in control and senescent cells according to the following procedures.
7.4.1.Calculate the average Ct for input RNA.
7.4.2.Subtract Ct of input RNA and log2(input dilution factor) from the Ct values obtained from the corresponding IP samples to calculate the normalized ΔCt.
7.4.3.Subtract the normalized ΔCt of control sample from the normalized ΔCt of both samples to calculate the ΔΔCt.
7.4.4.Determine the fold enrichment as 2^ (-ΔΔCt), thus normalizing the relative average fold enrichment of control sample to “1”.
Tips: To obtain the results with higher reliability, apply multiple non-specific IgG control antibodies and control primer pairs targeting m6A-negative sites. On one hand, include samples incubated with equal amount of IgG control antibodies when performing the immunoprecipitation step with m6A-specific antibody. On the other hand, include negative primers targeting the transcript segments without m6A modification when performing the qPCR step. Moreover, it is recommended that at least three independent experiments be done for each assay.
The RT-qPCR results are listed in Fig. 2B, which shows a decrease in m6A enrichment on MIS12 mRNA of senescent cells compared to control cells.
Figure 2. m6A modification on MIS12 mRNA in young and senescent cells. A. IGV views showed the m6A signals on MIS12 transcript in young and senescent cells. Peaks are represented as subtracted read densities (IP minus input). Black arrows indicate the forward and reverse primers. B. MeRIP-RT-qPCR was conducted to examine the m6A enrichment on MIS12 mRNA in young and senescent cells. Statistical analysis was performed by using GraphPad Prism (Student’s t-test).
Data are presented as the mean ± SEM. n = 6. ***, P < 0.001. These data are from a previously published study [14].
Troubleshooting
Potential issues and suggestions:
| Item | Description | Causes | Suggestions |
|---|---|---|---|
| I | Low m6A-enriched RNA is eluted after m6A IP and N6-methyladenosine elution | Low RNA input or significant loss during washing | Increase the RNA starting amount or reduce the times of washing |
| II | High background noise in the meRIP-seq data | Low m6A signal-to-background ratio | Use freshly prepared IP buffer, low/high salt buffer and the elution buffer |
Discussion
Epigenetic alterations and cellular senescence are believed to be critical hallmarks of aging, and these two processes can be targeted therapeutically to manage aging-associated disorders [2,4,10,22-26]. However, our understanding of the epitranscriptomic regulation of cellular senescence is still limited. Although more advanced m6A sequencing strategies with single-base resolutions have emerged [27,28], meRIP-seq remains a popular and straightforward approach for the analysis of m6A alterations across the entire transcriptome in aging and cellular senescence [14,21,29]. Here, we detailed a protocol for performing meRIP-seq and RT-qPCR validation in young (control) and senescent (treated) cells. We also included critical tips for key steps. Taking MIS12 as an example [14], we observed a reduced m6A level on its transcript in senescent cells relative to young cells, which was further validated by meRIP-RT-qPCR. For functional characterization of downstream genes, we recommend knockdown, knockout or overexpression experiments, complemented by senescence-associated phenotypic analyses, such as SA-β-gal staining, detection of senescence-associated gene expression, heterochromatin analysis, among others. [14,15,30,31]. Additionally, to test the necessity of m6A in the regulation of these processes, RNA methyltransferase/demethylase inhibitors, catalytically-inactive mutants, m6A site mutation or reader-related experiments are viable options [14,15,32-36].
Acknowledgements
We are grateful to Lei Bai engineer for her administrative assistance. This work was supported by the National Key Research and Development Program of China (2022YFA1103700, 2020YFA0804000, 2020YFA0112200, 2019YFA0110900, 2022YFA1103800, 2021YFF1201000), the National Natural Science Foundation of China (32100937, 81921006, 92149301, 92168201, 82173061, 82125011, 91949209, 92049304, 92049116, 32121001, 82192863), the Program of the Beijing Natural Science Foundation (Z190019), CAS Project for Young Scientists in Basic Research (YSBR-076, YSBR-012), New Cornerstone Science Foundation through the XPLORER PRIZE (2021-1045) CAS Special Research Assistant (SRA) Program.
References
- Di Micco R, Krizhanovsky V, Baker D, d'Adda di Fagagna F. Cellular senescence in ageing: from mechanisms to therapeutic opportunities. Nat Rev Mol Cell Biol. 2021;22(2):75-95. Epub 2020/12/18. https://doi.org/10.1038/s41580-020-00314-w PMID: 33328614
- López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: An expanding universe. Cell. 2023;186(2):243-78. Epub 2023/01/05. https://doi.org/10.1016/j.cell.2022.11.001 PMID: 36599349
- Gorgoulis V, Adams PD, Alimonti A, Bennett DC, Bischof O, Bishop C, et al. Cellular Senescence: Defining a Path Forward. Cell. 2019;179(4):813-27. Epub 2019/11/02. https://doi.org/10.1016/j.cell.2019.10.005 PMID: 31675495
- Liu B, Qu J, Zhang W, Izpisua Belmonte JC, Liu GH. A stem cell aging framework, from mechanisms to interventions. Cell Rep. 2022;41(3):111451. Epub 2022/10/20. https://doi.org/10.1016/j.celrep.2022.111451 PMID: 36261013
- Cai Y, Ji Z, Wang S, Zhang W, Qu J, Belmonte JC, et al. Genetic Enhancement: a New Avenue to Combat Aging-related Diseases. Life Med. 2022;1(3):307–18. https://doi.org/10.1093/lifemedi/lnac054
- Sun Y, Li Q, Kirkland JL. Targeting senescent cells for a healthier longevity: the roadmap for an era of global aging. Life Med. 2022;1(2):103–19. https://doi.org/10.1093/lifemedi/lnac030 PMID: 36699942
- Cai Y, Song W, Li J, Jing Y, Liang C, Zhang L, et al. The landscape of aging. Sci China Life Sci. 2022;65(12):2354-454. Epub 2022/09/07. https://doi.org/10.1007/s11427-022-2161-3 PMID: 36066811
- Yang Y, Li X, Zhang T, Xu D. RIP kinases and necroptosis in aging and aging-related diseases. LifeMed. 2022;1(1):2-20. https://doi.org/10.1093/lifemedi/lnac003
- Wu Z, Wang S, Belmonte JC, Zhang W, Qu J, Liu GH. Emerging role of RNA m6A modification in aging regulation. Curr Med (Cham). 2022;1(1):8. https://doi.org/10.1007/s44194-022-00009-8
- Wang K, Liu H, Hu Q, Wang L, Liu J, Zheng Z, et al. Epigenetic regulation of aging: implications for interventions of aging and diseases. Signal Transduct Target Ther. 2022;7(1):374. Epub 2022/11/07. https://doi.org/10.1038/s41392-022-01211-8 PMID: 36336680
- Roignant JY, Soller M. m(6)A in mRNA: An Ancient Mechanism for Fine-Tuning Gene Expression. Trends Genet. 2017;33(6):380-90. Epub 2017/05/14. https://doi.org/10.1016/j.tig.2017.04.003 PMID: 28499622
- Niu Y, Zhao X, Wu YS, Li MM, Wang XJ, Yang YG. N6-methyl-adenosine (m6A) in RNA: an old modification with a novel epigenetic function. Genomics Proteomics Bioinformatics. 2013;11(1):8-17. Epub 2013/03/05. https://doi.org/10.1016/j.gpb.2012.12.002 PMID: 23453015
- Deng X, Su R, Weng H, Huang H, Li Z, Chen J. RNA N(6)-methyladenosine modification in cancers: current status and perspectives. Cell Res. 2018;28(5):507-17. https://doi.org/10.1093/nar/gkaa816 PMID: 29686311
- Wu Z, Shi Y, Lu M, Song M, Yu Z, Wang J, et al. METTL3 counteracts premature aging via m6A-dependent stabilization of MIS12 mRNA. Nucleic Acids Res. 2020;48(19):11083-96. Epub 2020/10/10. https://doi.org/10.1093/nar/gkaa816 PMID: 33035345
- Wu Z, Lu M, Liu D, Shi Y, Ren J, Wang S, et al. m6A epitranscriptomic regulation of tissue homeostasis during primate aging. Nat Aging. 2023;3(6):705–21. https://doi.org/10.1038/s43587-023-00393-2 PMID: 37118553
- Li Q, Li X, Tang H, Jiang B, Dou Y, Gorospe M, et al. NSUN2-Mediated m5C Methylation and METTL3/METTL14-Mediated m6A Methylation Cooperatively Enhance p21 Translation. J Cell Biochem. 2017;118(9):2587-98. Epub 2017/03/02. https://doi.org/10.1002/jcb.25957 PMID: 28247949
- Dominissini D, Moshitch-Moshkovitz S, Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature. 2012;485(7397):2016. Epub 2012/05/12. https://doi.org/10.1038/nature11112 PMID: 22575960
- Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3' UTRs and near stop codons. Cell. 2012;149(7):1635-46. Epub 2012/05/23. https://doi.org/10.1016/j.cell.2012.05.003 PMID: 22608085
- Li X, Xiong X, Yi C. Epitranscriptome sequencing technologies: decoding RNA modifications. Nat Methods. 2016;14(1):23–31. https://doi.org/10.1038/nmeth.4110 PMID: 28032622
- Song J, Yi C. Chemical Modifications to RNA: A New Layer of Gene Expression Regulation. ACS Chem Biol. 2017;12(2):316-25. Epub 2017/01/05. https://doi.org/10.1021/acschembio.6b00960 PMID: 28051309
- Min KW, Zealy RW, Davila S, Fomin M, Cummings JC, Makowsky D, et al. Profiling of m6A RNA modifications identified an age-associated regulation of AGO2 mRNA stability. Aging Cell. 2018;17(3):e12753. Epub 2018/03/25. https://doi.org/10.1111/acel.12753 PMID: 29573145
- Zhang W, Qu J, Liu GH, Belmonte JC. The ageing epigenome and its rejuvenation. Nat Rev Mol Cell Biol. 2020;21(3):137–50. https://doi.org/10.1038/s41580-019-0204-5 PMID: 32020082
- Liu X, Liu Z, Wu Z, Ren J, Fan Y, Sun L, et al. Resurrection of endogenous retroviruses during aging reinforces senescence. Cell. 2023;186(2):287-304.e26. Epub 2023/01/08. https://doi.org/10.1016/j.cell.2022.12.017 PMID: 36610399
- Zhao D, Chen S. Failures at every level: breakdown of the epigenetic machinery of aging. LifeMed. 2022;1(2):81-3. https://doi.org/10.1093/lifemedi/lnac016.
- Zhu J, An Y, Wang X, Huang L, Kong W, Gao M, et al. The natural product rotundic acid treats both aging and obesity by inhibiting PTP1B. LifeMed. 2022;1(3):372-86. https://doi.org/10.1093/lifemedi/lnac044 PMID: 36811113
- Zhou S, Liu L, Lu X. Endogenous retroviruses make aging go viral. Life Med. 2023;2(1):lnad001. https://doi.org/10.1093/lifemedi/lnad001.
- Liu C, Sun H, Yi Y, Shen W, Li K, Xiao Y, et al. Absolute quantification of single-base m(6)A methylation in the mammalian transcriptome using GLORI. Nat Biotechnol. 2023;41(3):355-66. Epub 2022/10/28. https://doi.org/10.1038/s41587-022-01487-9 PMID: 36302990
- Hu L, Liu S, Peng Y, Ge R, Su R, Senevirathne C, et al. m(6)A RNA modifications are measured at single-base resolution across the mammalian transcriptome. Nat Biotechnol. 2022;40(8):1210-9. Epub 2022/03/16. https://doi.org/10.1038/s41587-022-01243-z PMID: 35288668
- Luo H, Liu W, Zhang Y, Yang Y, Jiang X, Wu S, et al. METTL3-mediated m(6)A modification regulates cell cycle progression of dental pulp stem cells. Stem Cell Res Ther. 2021;12(1):159. Epub 2021/03/03. https://doi.org/10.1186/s13287-021-02223-x PMID: 33648590
- Zhang J, Ao Y, Zhang Z, Mo Y, Peng L, Jiang Y, et al. Lamin A safeguards the m(6) A methylase METTL14 nuclear speckle reservoir to prevent cellular senescence. Aging Cell. 2020;19(10):e13215. Epub 2020/08/20. https://doi.org/10.1111/acel.13215 PMID: 32813328
- Zhang S, Wu Z, Shi Y, Wang S, Ren J, Yu Z, et al. FTO stabilizes MIS12 and counteracts senescence. Protein Cell. 2022; 13(12);954-60. https://doi.org/10.1007/s13238-022-00914-6 PMID: 35384602
- Yankova E, Blackaby W, Albertella M, Rak J, De Braekeleer E, Tsagkogeorga G, et al. Small-molecule inhibition of METTL3 as a strategy against myeloid leukaemia. Nature. 2021;593(7860):597-601. Epub 2021/04/27. https://doi.org/10.1038/s41586-021-03536-w PMID: 33902106
- Huang Y, Su R, Sheng Y, Dong L, Dong Z, Xu H, et al. Small-Molecule Targeting of Oncogenic FTO Demethylase in Acute Myeloid Leukemia. Cancer Cell. 2019;35(4):677-91.e10. Epub 2019/04/17. https://doi.org/10.1016/j.ccell.2019.03.006 PMID: 30991027
- Wei X, Huo Y, Pi J, Gao Y, Rao S, He M, et al. METTL3 preferentially enhances non-m(6)A translation of epigenetic factors and promotes tumourigenesis. Nat Cell Biol. 2022;24(8):1278-90. Epub 2022/08/05. https://doi.org/10.1038/s41556-022-00968-y PMID: 35927451
- Huang H, Weng H, Sun W, Qin X, Shi H, Wu H, et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol. 2018;20(3):285-95. https://doi.org/10.1038/s41556-018-0045-z PMID: 29476152
- Bai Y, Li K, Peng J, Yi C. m 6A modification: a new avenue for anti-cancer therapy. Life Med. 2023;2(1):Inad008. https://doi.org/10.1093/lifemedi/lnad008